
Регуляция была продемонстрирована на многих развивающихся системах. Однако в процессе развития организма эта способность утрачивается, когда определяется судьба его отдельных частей. Но даже в системах, где эта детерминированность имеет место на ранней стадии, например в эмбрионах насекомых, регуляция может осуществляться после повреждения яйца (рис. 1).
Результаты такого типа показывают, что развивающиеся системы движутся к морфологической цели и что они обладают некоторым свойством, которое определяет эту цель и позволяет им достичь ее, даже если части системы удалены и созданы препятствия для нормального хода развития.
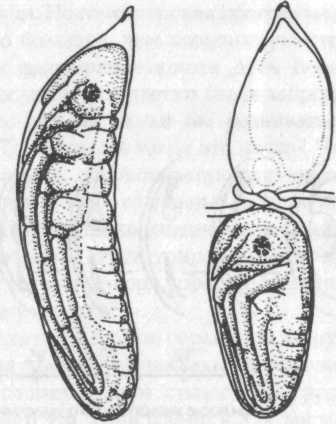
Рис. 1. Пример регуляции. Слева — нормальный эмбрион стрекозы Platycnemis pennipes. Справа — небольшой, но полноценный эмбрион, полученный из задней части яйца, перетянутого в середине вскоре после его откладывания (из Weiss, 1942)
Третья проблема — это регенерация, то есть способность организмов заменять или восстанавливать поврежденные структуры. Растения обнаруживают удивительно широкий диапазон возможностей регенерации, и то же можно сказать о низших животных:
например, если плоского червя разрезать на несколько кусочков, из каждого может вырасти целый червь. Даже многие позвоночные обладают поразительными способностями к регенерации: например, если хирургическим путем удалить линзу из глаза тритона, из края радужной оболочки вырастает новая линза (рис. 2); при нормальном эмбриональном развитии линза образуется совершенно другим путем, из клеток кожи.