
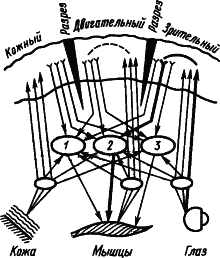
Рис. 9. Множественное замыкание временных связей в коре (показано пунктиром), которым не препятствуют ее разрезы (по А.Б. Когану):
1, 2, 3 — центральные механизмы оборонительных, пищевых и ориентировочных реакций соответственно; путь условного пищевого рефлекса на световой сигнал показан жирными линиями
Как показали многочисленные исследования, участие подкорковых структур в процессах высшей нервной деятельности не ограничивается регуляторной ролью ретикулярной формации среднего мозга и лимбических структур. Ведь уже на подкорковом уровне происходят анализ и синтез действующих раздражителей и оценка их биологического значения, в значительной мере определяющая характер образующихся связей с сигналом. Использование показателей формирования наиболее коротких путей, по которым сигнал достигает разных подкорковых структур мозга, выявило наиболее выраженное участие в процессах обучения задних отделов таламуса и поля CA3 гиппокампа. Роль гиппокампа в явлениях памяти подтверждается многими фактами. Наконец, нет оснований предполагать, что способность к примитивной замыкательной деятельности структур мозга, которая приобреталась в эволюции, когда они были ведущими, теперь у них полностью исчезла, когда эта функция перешла к новой коре.
Таким образом, корково-подкорковые отношения определяются регулированием функционального состояния коры активирующей системой — ретикулярной формацией среднего мозга и тормозной системой неспецифических ядер таламуса, а также возможным участием в образовании примитивных временных связей на нижнем уровне сложных иерархических механизмов высшей нервной деятельности.
Межполушарные отношения. Каким образом полушария головного мозга, являющиеся парным органом, участвуют в процессах образования условных связей? Ответ на этот вопрос был получен в опытах на животных, которым была сделана операция «расщепления» мозга путем перерезки мозолистого тела и передней комиссуры, а также продольного разделения перекреста зрительных нервов (рис. 10).